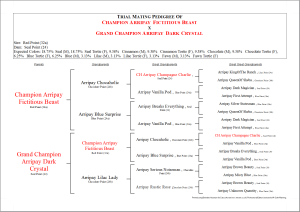
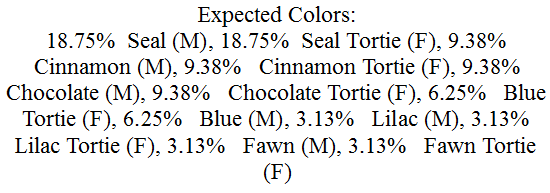
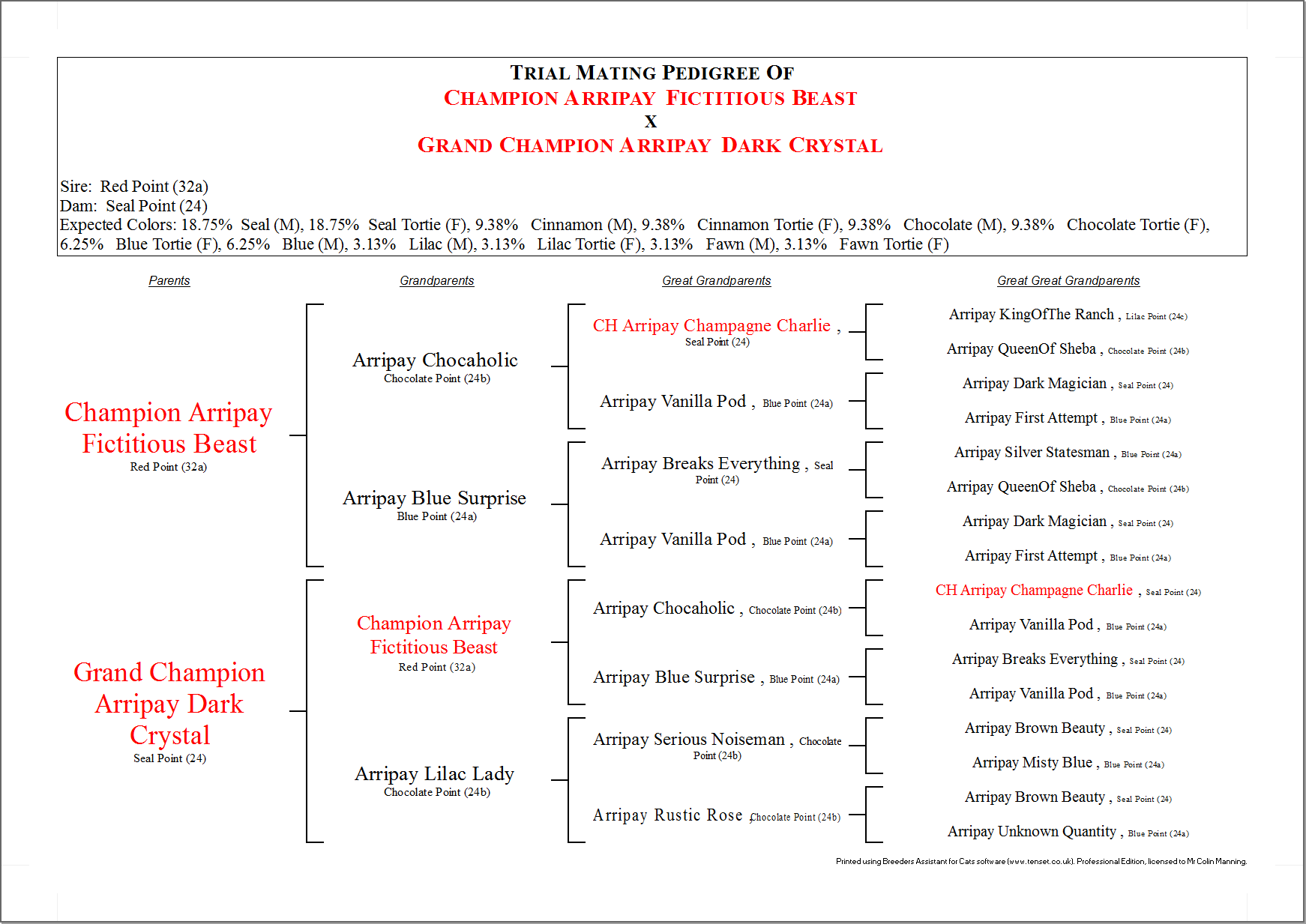
Color Genes of the Pedigree Cat
Much of what is given here assumes you are familiar with basic Mendelian genetics - i.e. you understand what is meant by the terms 'gene', 'allele', 'recessive', 'phenotype' etc.
The relevant genes can be divided into the following groups:
- Color Genes: genes that affect the pigment color of hairs.
- Pattern Genes: genes that affect the distribution of a particular color.
- Breed Genes: genes that generally typify all variants of a given breed, but excluding color and pattern genes. These broadly divide into coat type genes: longhair, wirehair etc., and some genes that are really congenital defects but which are considered normal characteristics for some breeds e.g. Polydactyly.
Breed vs Genetic Terminology
Different terms are sometimes used for the same genetic colors, depending on breed and sometimes country too. This can really confuse matters. E.g. a cat that is genetically 'black' (B- D- o/oo aa) is also known as 'Brown' in Burmese, 'Seal' in Siamese, etc.
This article will use the 'genetic' color descriptions.
Melanin, Agouti, and Red
This is quite a complex topic but its understanding is fundamental to understanding the basic color genetics of the cat. Many of the other genes are simple recessives that have easy-to-understand effects.
Melanin is the substance or pigment that gives a cat's hair its color. There are two distinct types of melanin in the cat - as in many mammals - eumelanin and phaeomelanin. Eumelanin is, in the absence of other modifying genes (see below), black or dark brown. Phaeomelanin is, in its unmodified form, a yellowish color though cat breeders generally call it red.
Melanin is produced by cells called melanocytes. These are found in the skin, hair bulbs (from which the hairs grow) and other places.
Melanocytes in the skin are sensitive to UV radiation and normally produce eumelanin. They are responsible e.g. for tanning in humans, and for darker exposed skin e.g. nose leather in furred mammals.
Melanocytes within the hair follicles cause melanin to be added to the hair as it grows. However, melanin is not added at a constant 'rate'. At the very tip of the hair, (eu)melanin production is usually most intense, resulting in the darker tip that's frequently seen.
A protein called the Agouti protein has a major effect on the 'injection' of melanin into the growing hair. This is widespread in many mammals, though not in humans. The Agouti protein causes a banding effect on the hair: it causes a fairly sudden change from the production of eumelanin (black/brown pigment) to phaeomelanin (red/yellow pigment). The result is the ticked, agouti appearance as typified by the wild rabbit. The term 'Agouti' actually refers to a South American rodent that exemplifies this type of hair.
In the domestic cat, however, there are three separate factors that between them conspire to generate the 'primary' colors and patterns:
- the non-agouti ('self') mutant form of Agouti protein;
- tabby patterning; and
- the sex-linked Red (O) gene.
Non-Agouti
There is a mutant form of the Agouti protein, resulting from a recessive gene (symbol a). When a cat is homozygous for a (i.e., aa), the Agouti protein in the melanocytes is replaced by a 'faulty' form that has a much lesser (though not zero) inhibitory effect on eumelanin. This results in hairs that contain eumelanin throughout, giving rise to solid ('self') colors.
Tabby Patterns
Tabby patterns (mackerel, blotched, spotted) are controlled by genes that cause the amount of agouti protein (or 'faulty' agouti protein, if the cat is aa) in different areas of the cat's coat to be greatly reduced. It has the effect of preventing the normal shift from eumelanin to phaeomelanin. Thus in a tabby cat the hairs in the black parts where the tabby pattern shows through are eumelanin throughout, whereas in the other areas the typical agouti ticking is seen.
Its worth noting that the effect of the tabby patterning genes on the hairs is similar, but not identical, to the non-agouti gene. This can be seen in the self colored cat that still shows tabby patterns or barring.
The various tabby genes are covered in more detail below.
Sex-linked Red
This is a gene with symbol O and is probably the most interesting gene affecting cat color/pattern from a genetic point of view.
Its effect is to replace all the eumelanin in the growing hairs with phaeomelanin. I.e, the hairs exhibit red-based rather than black-based colors.
What makes it more interesting though is that:
- The O gene is sex-linked on the X chromosome, and
- In the heterozygote (female - Oo), it causes the Tortoiseshell pattern. So male cats are either O (red) or o (black), whereas females can be OO (red), Oo (tortoiseshell) or oo (black). [For simplicity this ignores rare cases of tortie males - these result from abnormal construction of the sex chromosomes].
Red with Tabby: phaeomelanin pigment is affected by both forms of the agouti protein to more or less the same extent. The important point is that it is affected. And for this reason, tabby patterns do show up in the phaeomelanin-based coat of the red cat, because these patterns are a result of reduced levels of the agouti/non-agouti protein. The tabby markings show as a richer form of the red-based color, because the phaeomelanin has been inhibited to a lesser degree.
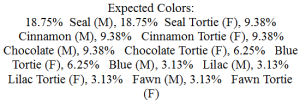
Tortoiseshell: a patchwork effect where some parts of the coat have eumelanin and the others phaeomelanin. The eumelanistic parts are affected by all the normal modifying genes that are active on eumelanin: B, D, Dm and in the presence of normal Agouti (A-) will display tabby markings prominently. The phaeomelanistic parts are affected by D and Dm in the usual way and tabby markings can continue through from the eumelanistic parts, this time showing as a richer red. The 'richness' of red coloration is also affected by what are known as 'rufous polygenes' (see below).
Color Genes
These are the genes that affect the pigment colors (as opposed to pigment distribution or pattern - either within the hairs or over the body).
| Gene | Effect | ||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chocolate - B gene |
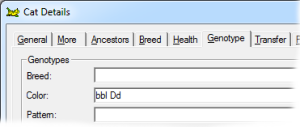
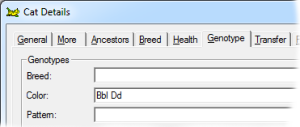
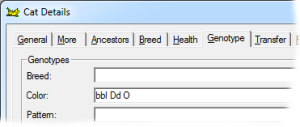
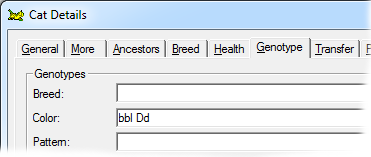
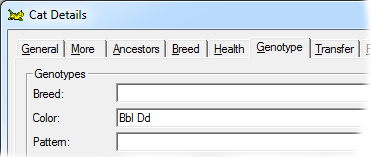
This gene has a lightening effect on eumelanin only. I.e, it has no effect on red-based colors. There are three alleles for this gene, with symbols B, b and bl respectively, in order of dominance. When B is present (BB, Bb or Bbl) the brown/black eumelanin is its normal, unlightened, color. If a cat is bb or bbl the brown is lightened to chocolate. When a cat is homozygous for bl the color is further lightened to cinnamon. |
||||||||||||||||||||||||||||||||||||||||
| Blue Dilution - D gene |
This recessive gene has a diluting effect on both eumelanin and phaeomelanin. When present in the homozygous recessive form (dd) it dilutes brown eumelanin to blue, and red to cream. |
||||||||||||||||||||||||||||||||||||||||
| Blue Modifier - Dm gene |
This is a dominant gene that only affects the phenotype in cats that are already homozygous for blue dilution (dd). It has a browning effect on the otherwise diluted color. Blue becomes Caramel, and Cream becomes Apricot. |
||||||||||||||||||||||||||||||||||||||||
| Combinations of B, D, and Dm in Eumelanistic Coats |
The effects of these 3 genes combine to form the full range of eumelanistic ('black-based') colors:
|
||||||||||||||||||||||||||||||||||||||||
| Combinations of D and Dm in Phaeomelanistic Coats |
The effects of these 2 genes combine to form the full range of phaeomelanistic ('red-based') colors:
|
||||||||||||||||||||||||||||||||||||||||
| Red - O gene |
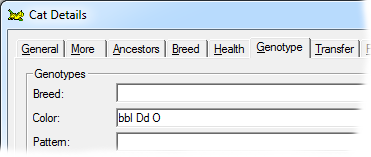
A sex linked trait. Males can be O or o; females can be OO, Oo or oo. The effect of O is to replace all the eumelanin in the hairs with phaeomelanin, however in the heterozygote female Oo, this gives rise to the tortoiseshell pattern of intermingled areas of eumelanin pigment (black-based) and phaeomelanin (red-based). |
||||||||||||||||||||||||||||||||||||||||
| Albino - C gene |
This gene affects the intensity of melanin production. The normal or dominant form, C, is what might be termed 'full color'. There are however various incompletely dominant mutant alleles possible for this locus, with varying effects on color intensity. These mutant forms are temperature sensitive - the higher the temperature, the more effective they are (i.e., the lighter the color). In order of decreasing dominance the alleles are C, cb, cs, ca and c. C is the wild-type or full color expression and is dominant to all other alleles at this locus. cb is the 'Burmese' factor; it causes a slight lightening of black (eumelanin) to brown or dark sepia, and of orange (phaeomelanin) to a yellow color, and is only slightly temperature sensitive. A Brown Burmese (US: Sable) will be slightly darker at the points but not markedly as in the Siamese. cs is the 'Siamese' factor; it has a much greater lightening effect than the Burmese allele and appears more temperature sensitive, with the points being considerably darker than the body. cb is incompletely dominant over cs; the heterozygote (cbcs) gives a phenotype intermediate between Burmese and Siamese, known as Tonkinese. The most recessive form is c, also known as pink-eyed albino. In the homozygote cc this causes complete absence of any pigment in both the fur and eyes: white fur and pink eyes (the pink color is simply the color of the blood). This is comparatively rare in the pedigreed cat. It is possible there is another mutation between c and cs, sometimes symbolised as ca, that results in there being a very small amount of pigment in the eye giving rise to pale blue eyes, but still none in the coat (white). Neither c nor the possible pale-blue-eyed mutant version of it should be confused with a quite separate mechanism that can also produce a pure white coat: the dominant white (see below). |
||||||||||||||||||||||||||||||||||||||||
| Dominant White - W gene |
Entirely white coats in cats are produced by the pink-eyed albino genotype (cc), possibly by the pale-blue-eyed albino (caca or cac) or more likely by a quite separate mechanism known as dominant white. This gene has symbol W. When present (either Ww or WW) it causes an entirely white coat, overiding all other color genes. Dominant white also affects eye color, but not consistently. Many dominant white cats are blue eyed, others are odd eyed. White cats are also more likely to suffer from deafness than non-white cats. |
||||||||||||||||||||||||||||||||||||||||
| Bengal Glitter - Gl gene |
This is a simple recessive gene which causes a glittering of the coat in Bengals. Breeders Assistant use the symbol Gl for this, glittered being expressed in the homozygote (glgl). |
||||||||||||||||||||||||||||||||||||||||
| Rufousing - polygenic |
The 'richness' of phaeomelanin pigment is affected by what are known as 'rufous polygenes'. Breeders of red cats can selectively breed for these richer red tones e.g. as seen in the red tabby, and 'Usual' Abysinnian. In some breeds this effect is undesirable, where it is sometimes called tarnishing e.g. in cats that are heterozygous for the silvering gene I (see below). |
Pattern Genes
The most important genes that affect pattern are the various tabby genes. Many breeders have, understandably, associated tabby with the Agouti (A) gene because of the effect of the non-agouti recessive self form (aa), however the tabby patterns themselves are not controlled by the A gene; what the A gene controls is the degree to which the tabby patterns will be visible (either highly, especially in an eumelanistic coat, or hardly if at all).
| Gene | Effect |
|---|---|
| Agouti - A |
The Agouti gene controls whether the cat has the 'normal' agouti protein (A-) or the mutant defective form of it (aa). As discussed above, the normal form causes the hairs to have a characteristic band of phaeomelanin (yellow) after the tip, which is normally eumelanin based. In a cat that is not red (male o or female oo), or in the 'black' parts of a tortoiseshell (female Oo), the state of the Agouti gene is what affects whether any tabby patterning will be highly visible (A-) or hardly visible if at all (aa). |
| Tabby Genes - Mc, Ta, Sp |
There are several forms of tabby pattern in the cat: Mackerel and blotched patterns are controlled by the Mc gene, with mackerel being the dominant form (Mc-) and blotched being the recessive form (mcmc). Ticked tabby is controlled by a gene with symbol Ta. The dominant allele Ta is dominant over mackerel and blotched patterns in the homozygote (TaTa), but only partially so in the heterozygote (Tat). E.g. an Abysinnian that's heterozygous for Ta may well have faint tabby markings - whether they are mackerel or blotched depends on whether or not Mc is present. Ticked tabby also appears to mask spotted tabby (Sp-). The ticked form of tabby is characterised by the coat not having the distinctive barring of either mackerel and blotched; the appearance is that of a more universally ticked coat. For this reason, many breeds where any kind of tabby barring are undesirable are often TaTa for the ticked tabby gene, even though the breed isn't considered to be a 'ticked' breed. E.g. Burmese are ideally TaTa to reduce tabby markings whereas those with slight barring are more likely to be Tat or tt. Mackerel and ticked tabbies are always less pigmented than blotched, which is why solid red-based cats are often bred to be mcmc as this increases the richness. Spotted tabby, Sp, is a dominant modifier of the blotched/mackerel tabby pattern. It causes the pattern to break up into spots, though the underlying pattern of blotched/mackerel is still discernable. In many breeds where spotted tabbies are found, an underlying blotched pattern is preferable. It should be noted, this is not the same kind of spotting as is found in breeds such as the Bengal - the spotting in those breeds may be termed non-domestic spotting and the spots tend to come in horizontal lines. |
| Silvering - I |
This is an incompletely dominant modifier gene that inhibits the addition of melanin into growing hairs. This causes hairs that are white but with color at the very tip, an effect also known as 'silvering'. This gene is responsible for the silver tabby and smokes. The mutant form has a greater effect on phaeomelanin(red) than eumelanin(black). |
| White Spotting (Piebaldism) - S |
The S gene is responsible for piebald or white spotting, and is the gene that causes Bi-Color, Tri-Color and Van patterns. It is an incomplete dominant. The cat that is homozygous for S will generally have more extensive white spotting than the heterozygote. The expression of S varies considerably, and there is doubt over whether there are in fact other genes at play, or other alleles at the S locus. In other mammals, e.g. dogs, there are thought to be additional alleles present. As with cats there are doubts in the dog world over the precise nature of the alleles of this gene and their interactions. Cats exhibiting extensive white spotting are more likely to suffer from deafness than non-white cats. |
Breed Genes
So far this article has only considered genes that affect coat color and pattern. There are of course many, many others. Often these genes will always be the same 'state' for a given breed and are characteristic of that breed. Some of the more well established ones are listed here.
Most of these genes are either genes that affect hair length or wavyness ('Rex'), or they are genes that are, in reality, congenital deformities but which are nevertheless recognised as breeds in their own right, particularly in the US.
| Gene | Effect |
|---|---|
| Longhair - L |
The recessive gene that gives rise to the longhaired breeds (ll). Sometimes the heterozygote may have a clearly different phenotype to the homozygote. |
| Devon Rex - Re |
A recessive gene that gives rise to the Devon Rex (rere or rehr) and Sphynx (hrhr). The non-Devon-rexed cat is Re-. |
| Cornish Rex - R |
Another recessive gene responsible for the Cornish Rex (rr) coat. A third allele is also present at this locus for the German Rex, recessive to r (rgrg). The non-Cornish-rexed cat is R-. |
| Oregon Rex - Ro |
Another recessive wavyness gene giving rise to the Oregon Rex coat (roro). This gene may no longer be present in the cat population. |
| Selkirk Rex - Se |
This is a dominant mutation giving rise to the Selkirk Rex coat type (Se-). |
| Wirehair - Wh |
This dominant gene with incomplete penetrance gives rise to wirehair coat (Wh-). |
| Manx Tailed - M |
A dominant mutation that causes the spinal deformities characterised by the Manx cat (Mm). Lethal in the homozygote (MM). Polygenes control the length of the 'tail' - varying from no tail at all to a 'longie'. |
| Munchkin (Hypochondroplastic Dwarfism) - Mk |
A dominant form of dwarfism characterised by the Munchkin breed (Mk-). Causes shortened long bones in all legs, but unlike a related form of dwarfism, Achondroplastic Dwarfism, it does not affect head size. |
| Curled Ears - Cu |
A dominant mutation giving rise to a cosmetic backward curling of the ears as exemplified by the American Curl breed. |
| Japanese Bobtail - Jb |
A dominant mutation with incomplete penetrance causing a shortened tail as found in the Japanese Bobtail breed. Unlike the Manx mutation, this is not crippling and does not cause deformation of the spine. |
| Folded Ears - Fd |
A dominant mutation with incomplete penetrance causing a forward bending of the ears as in the Scottish Fold breed. In the homozygous form the cat may also suffer crippling complications such as swollen feet and a thickened tail. |
| Polydactyly - Pd |
A dominant mutation with variable expression giving rise to extra toes (Pd-). |
A Comparison With Dogs
It is perhaps interesting to compare cat color/pattern genetics with dog color genetics. For there are many similarities, and yet also some interesting differences.
The most striking difference is the gene that enables phaeomelanin (red/yellow). In dogs - and many other mammals for that matter - there is a gene called the extension gene, symbol E. The term 'extension' refers to the degree of extension of eumelanin across the coat. In the non-extended recessive form, the coat is entirely phaeomelanin based, sometimes called recessive red or recessive yellow by dog breeders. In the domestic cat however there is the sex-linked red gene (O).
The chocolate gene (B) is present in dogs, and has exactly the same effect, though the lighter phase bl allele does not seem to occur.
The blue dilution gene (D) also occurs, and works in the exact same way. Unlike cats however the recessive blue form (dd) is completely missing in many breeds, as is the dilute modifier gene Dm.
Piebaldism (white spotting) - the S gene - is also present in dogs, where as with cats aspects of the operation of this gene are not understood. It likewise is associated with deafness in dogs just as it is in cats.
One area of controversy in dogs is the inheritance of black. There seems to be two types: dominant black and recessive black. There is general agreement about the recessive form, in that it is at the bottom of the Agouti series (aa) and in that it is quite consistent with black-based solid colored cats. It is generally also agreed that there is a dominant form of black in many breeds of dog, and traditionally this has been put at the head of the Agouti series (As). However, this flies in the face of the behaviour of the Agouti series in almost all other mammals, none of which has a dominant phaeomelanin suppressor like As at the top of the series. Newer research indicates that 'dominant black' in dogs does not belong in the Agouti series at all but is a separate dominant gene (symbol K); the K series is also postulated as controlling brindle, with the brindle allele being intermediate between dominant black (K) and 'normal' (k). Robinson 4th Edn. mentions that it was thought at one time that there might be a dominant form of black in cats: "Tjebbes in 1924 published a report on crosses between Siamese and a tabby and indicated that the Siamese transmitted an epistatic gene that obscured the tabby pattern". However, there was no further breeding data to substantiate the claim, and perhaps even if correct the gene doesn't appear to be present in the present day pedigreed cat gene pool. It is perhaps interesting to speculate whether this may in fact be the same as the K series of current thinking in dogs.