Color Genes of Pedigree Dogs
This article assumes you understand Mendelian genetics - i.e. you know what is meant by the terms 'gene', 'allele', 'phenotype' etc.
The genes of interest can be broadly classified as either:
- Color Genes: genes that affect the pigment color of hairs; OR
- Pattern Genes: genes that affect the distribution of a particular color.
Breed vs Genetic Terminology
Different terms are sometimes used for the same genetic colors, depending on breed and sometimes country too. This can really confuse matters. E.g. a dog that is genetically 'recessive red' (ee) is known as yellow in some breeds. This article will try to stick to the 'genetic' color descriptions.
Melanin, Agouti, and Red
This is quite a complex topic but its understanding is fundamental to understanding the basic color genetics of the dog. Many of the other genes are simple recessives that have easy-to-understand effects.
Melanin is the substance or pigment that gives a dog's hair its color.
There are two distinct types of melanin in the dog - as in many mammals - eumelanin and phaeomelanin. Eumelanin is, in the absence of other modifying genes (see below), black or dark brown. Phaeomelanin is, in its unmodified form, a yellowish color.
Melanin is produced by cells called melanocytes. These are found in the skin, hair bulbs (from which the hairs grow) and other places.
Melanocytes in the skin are sensitive to UV radiation and normally produce eumelanin. They are responsible e.g. for tanning in humans, and for darker exposed skin e.g. nose leather in furred mammals.
Melanocytes within the hair follicles cause melanin to be added to the hair as it grows. However, melanin is not added at a constant 'rate'. At the very tip of the hair, (eu)melanin production is usually most intense, resulting in the darker tip that's frequently seen.
A protein called the Agouti protein has a major effect on the 'injection' of melanin into the growing hair. This is widespread in many mammals, though not in humans. The Agouti protein causes a banding effect on the hair: it causes a fairly sudden change from the production of eumelanin (black/brown pigment) to phaeomelanin (red/yellow pigment). The result is the agouti appearance as typified by the wild rabbit. The term 'Agouti' actually refers to a South American rodent that exemplifies this type of hair.
NOTE: It is worth mentioning at this point that there remain areas of considerable doubt about the exact genes/alleles that affect certain aspects of coat color in dogs. The main ones surround the proposed 'dominant black' (As) at the top of the Agouti series, and in the E series there are questionmarks over Em (black mask) and Ebr (brindle).
In the past it was thought that with the A and E series, you had:
1. Agouti Series (A): As, ay, aw, as, at, a
2. Extension Series (E): Em, E, Ebr, e
But newer research indicates that 'dominant black' As may not belong in the Agouti series at all - and this would be consistent with other mammals that also share the Agouti series, none of which has a dominant phaeomelanin suppressor like As at the top of the series. Research also indicates that brindle (Ebr in the older system) and 'dominant black' form a separate series of at least 3 alleles, which can be called the K series (for blacK); this includes both dominant black and brindle.
The present article adopts this later model.
| Gene | Effect |
|---|---|
| The Agouti Gene - A |
The Agouti locus controls the formation of the Agouti protein, that in turn is one of the mechanisms that controls the replacement of eumelanin with phaeomelanin in the growing hair. The alleles of the Agouti locus can affect not just whether or not the eumelanin->phaeomelanin shift occurs, but also where on the dog's body this happens. The probable alleles at the Agouti locus, in order of decreasing dominance, are: Ay, aw, as, at and a. Dominant Black. Dogs certainly do have a dominant form of black that is indeed very dominant: completely obliterating all formation of phaeomelanin pigment. Traditionally, dominant black has been placed at the head of the Agouti series (symbol As). It is now believed to be part of a separate series (the K series - see below) and not at the Agouti locus at all. This is in keeping with the operation of the Agouti locus in all other mammals that have it: increasing dominance of Agouti locus alleles results in increasing production of phaeomelanin without exception. It is mentioned here simply because it has long been thought, mistakenly, to be part of the Agouti locus. So at the top of the Agouti series is Ay, Sable - also known as 'dominant yellow' or 'golden sable'. This results in an essentially phaeomelanic phenotype, but the hair tips are eumelanin (black). The extent of the eumelanin tip varies considerably from lighter sables (where just the ear tips are black) to darker sables - where much of the body is dark. It is possible that Ay is not completely dominant over the lower Agouti series alleles, with an Ay heterozygote e.g. Ayat having a darker body. AyAy may be called 'clear red' whereas Ayat can be 'sabled red'. Sable is a very common color in many breeds of dog, e.g. German Shepherd. Next is aw, 'wolf' color. This is like Ay but the tan is replaced with a pale gray/cream color and the hairs usually have several bands of light and dark color, not just the black tip of sable. Seen in the Keeshond, Siberian and Norwegian Elkhound. Moving down the series the next allele is as, 'saddle tan'. This is somewhat like the black+tan allele (below), except that eumelanin is restricted to the back and side regions, hence the name 'saddle'. It is also possible that this is due to another gene interacting with at/at genotypes. Allele at, 'black+tan' is next. This is a primarily black dog but with tan (phaeomelanin) markings around the eyes, muzzle, chest, stomach and lower legs. Commonly seen in hounds, Dobermann's and Rottweilers. Finally, at the bottom of the Agouti series is recessive black, symbol a. When a dog is homozygous for recessive black (aa), it will have no phaeomelanin in its coat (unless it is also ee, which is epistatic to the Agouti series - see below). Examples of breeds that exhibit recessive black are German Shepherd and Shetland Sheepdog. Whilst some breeders discount the presence of a recessive black at the bottom of the Agouti series this is consistent with the behaviour of this locus in many other mammals. The existence of all these alleles in the Agouti series is not certain, nor is the precise order of dominance of the intermediate alleles aw as and at. |
| The Extension Gene - E |
This refers to the extension of eumelanin over the dog's body. The dominant form, E, is known as normal extension. The recessive form, e, is called non-extension. When a dog is homozygous for non-extension (ee), its coat will be entirely phaeomelanin based - i.e. red/yellow. For this reason this is sometimes termed recessive red or recessive yellow, to distinguish it from the sable reds/yellows generated by the Ay allele at the Agouti locus. The extension locus is shared by many mammals, e.g. horses, and operates in a very similar way to this description. In the past it had been proposed that there were additional alleles present at the E locus for black mask and brindle, but this is not borne out by breeding data. |
| The (Dominant) Black Gene - K |
This relative newcomer to the traditional models of dog color genetics codes for both dominant black and brindle in decreasing order of dominance: K=dominant blacK, kbr=brindle, k='normal'. So a dog that is KK or Kkbr is dominant black, kbrkbr or kbrk is brindled, and kk is 'normal'. Brindling is the presence of 'stripes' of eumelanin-based hairs in areas that are otherwise phaeomelanin based. When a dog is brindled the color of the eumelanin stripes may be modified by the normal genes that affect eumelanin (B and D, see below). Dominant black (K) is epistatic to whatever is found at the Agouti locus, however it in turn is overridden by ee at the E locus. |
Bringing it Together
It appears that the three loci E, K and A act together as follows:
- If a dog is ee at the E locus, its coat will be entirely phaeomelanin based (red/yellow);
- Otherwise (it is Ee or EE at the E locus), if it is K- at the K locus its coat will be entirely eumelanin based (dominant black);
- Otherwise (it is Ee or EE at the E locus), if it is kbrkbr or kbrk at the K locus it will be brindled with the color of the phaeomelanin part of the brindling in turn affected by the Agouti alleles present;
- Otherwise (it is Ee or EE at the E locus, and it is kk at the K locus), the distribution of eumelanin and phaeomelanin will be determined solely by the Agouti alleles present.
Other Genes
| Gene | Effect | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chocolate - B gene |
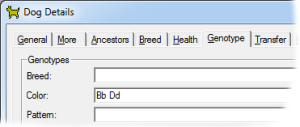
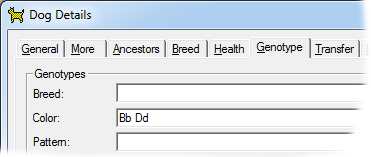
This gene has a lightening effect on eumelanin only. I.e, it has no effect on red-based colors. In the dog there are two alleles for this gene, with symbols B and b respectively. When B is present (BB or Bb) the brown/black eumelanin is its normal, unlightened, color. But when a dog is bb the brown is lightened to Chocolate. |
|||||||||||||||
| Blue Dilution - D gene |
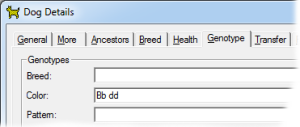
This recessive gene has a diluting effect on both eumelanin and phaeomelanin. When present in the homozygous recessive form (dd) it dilutes brown eumelanin to blue, and red to cream. |
|||||||||||||||
| Combinations of B and D in Eumelanistic Coats |
The effects of these 2 genes combine to form a range of 4 eumelanistic ('black-based') colors:
|
|||||||||||||||
| Albino - C gene |
This gene affects the intensity of melanin production in the coat hairs. The normal or dominant form, C, is what might be termed 'full color'. There are however various incompletely dominant mutant alleles postulated for this locus, with varying effects on color intensity. These mutant forms are temperature sensitive - the higher the temperature, the more effective they are (i.e, the lighter the color). Almost all dogs are CC at this locus - full color. The lower series alleles that have been suggested include, in order of decreasing dominance, cch, ce, cb and c. The first, cch, is chinchilla. This lightens most or all of the phaemelanin with little or no effect on eumelanin. E.g. it turns black+tan to black+silver. The next allele, ce, is 'extreme dilution'. Causes tan to become almost white. It is thought that the white labrador might be ce with another, lower, C series allele. The ce allele may be responsible for producing white hair in other breeds of dogs, like the West Highland White Terrier, while allowing full expression of dark nose and eye pigment. Next down the series is cb, or blue-eyed albino. This is an entirely white coat but with a very small amount of residual pigment in the eyes, giving pale blue eyes. This is known as ca in cats. Can be called platinum or silver. The most recessive allele in the series is c, true pink-eyed albino. This doesn't seem to occur in dogs. |
|||||||||||||||
| Graying - G gene |
This is a dominant mutant gene that causes the dog to gray with age - pigmented hairs are progressively replaced with unpigmented hairs. |
|||||||||||||||
| Super-Extension - Se |
This dominant gene controls the expression of a black mask. Traditional models of dog color genetics placed black mask in the Extension (E) series. However it seems that a more accurate model is to place it at a separate locus. Alternate literature has used the symbol Ma for this. Most breeds do not exhibit black mask, and are therefore sese for this locus. Breeds that have black mask (Se-) include the Mastiff, Pug and Belgian sheepdog. |
|||||||||||||||
| White Spotting - S |
The piebalding or 'white spotting' gene is common to many mammals e.g. cats, not just dogs. And, as in cats, it is not really fully understood. In dogs it is thought there are four alleles, in decreasing order of dominance: S, si, sp and sw. The S series alleles appear to be incompletely dominant, further complicating matters. E.g. Ssw will be similar to sisi in appearance. The degree of white spotting is also affected by modifiers. E.g when merle (M) is also present, the white spotting has a greater effect (more white). The most dominant allele, S, means 'solid color'. Most dogs that are homozygous for S (i.e. SS) have no white hair at all, or possible a tiny amount e.g. a white tail tip, though this can be considered a fault in breeds that are normally SS. The next allele, si, mean 'irish spotting'. This involves white spotting on most parts of the coat, but not crossing the back. Moving down the series is sp, 'piebald'. The white is more extensive than irish spotting, and often crosses the back. The most recessive allele in the series is sw, or 'extreme white piebald'. A dog that is homozygous for sw will be almost entirely white, e.g. the Dalmatian. Breeds that exhibit white spotting use varying terminology for the differing degrees of white. E.g. in herding dogs the terms 'normal white pattern' (sisi), 'white-factored' (sisw) and 'color headed' (swsw) are seen. In Boxers there is the term 'flashy white' (Ssw - si is not thought to exist in the Boxer). The term 'Mantle' is applied to a Great Dane that is sisw. Dogs exhibiting extensive white spotting are more likely to suffer from deafness than non-white dogs, e.g. Dalmatians. |
|||||||||||||||
| Ticked - T |
This dominant mutation causes the presence of color in those areas that have been made white by the effect of alleles in the white spotting (S) series. An extreme example of ticking is the Dalmatian. |
|||||||||||||||
| Merle - M |
This is an incomplete dominant that causes 'merling' - patchy dilution e.g. black becoming patched with gray ('blue merled') or sable becoming sable merle. Merling has little or no effect on phaeomelanin. The M allele is not found in all breeds; in fact most don't have it. Examples of breeds that display merling include the Australian Shepherd and Dachshund (where it is called 'dapple'). Merling also affects the eye color. Most merled dogs are actually the heterozygote, Mm. There is good reason for this; there are serious health problems associated with the homoygote MM, including tiny eyes (microopthalmic) or even entirely missing eyes (anopthamlic). The homozygote is also known as a 'double merle'. They are often predominantly white, hence their alternate name 'defective white'. When breeding merles, many reputable breeders avoid breeding Mm to Mm; they generally go for Mm X mm and get 50% single merles (Mm) and 50% not. |
A Comparison With Cats
It is perhaps interesting to compare dog color/pattern genetics with cat color genetics. For there are many similarities, and yet also some interesting differences.
The most striking difference is that cats do not have the Extension (E) gene. But they do have a fascinating alternative - the so-called Red gene (O). The effect of the O allele is to cause eumelanin to be replaced with phaeomelanin. But what makes O particularly interesting is that it is sex-linked. Males can be O or o, but females can be OO, Oo or oo. And the heterozygote, Oo is also a fascinating case because it is this that causes the Tortoiseshell pattern - where the cat's coat is made up of a patchwork of eumelanistic areas (black based) and phaeomelanistic areas (red based).
Cats also do not seem to have a dominant form of black, nor do they exhibit brindling.
Cats do have the Agouti series - though there is thought to be just 2 alleles present which correspond roughly to Ay and a in the dog.
Eumelanistic colors are affected by blue dilution (D gene) and chocolate (B) just as they are in the dog, but with some additions. The B gene in the cat has a third, more recessive allele, symbol bl, which gives rises to a lighter phase of chocolate (called cinnamon). Secondly, there is a dominant modifier to the blue dilution gene that causes a brownish tinge to cats that are homozygous for blue dilution (dd). Blue dilution has the same effect on phaeomelanin in cats as it does in dogs.
Piebaldism (white spotting) - the S gene - is also present in cats, and as with dogs aspects of the operation of this gene are not understood. It likewise is associated with deafness in cats just as it is in dogs. Cat breeders generally consider there to be 2 alleles (S and s) in the series, with S being incompletely dominant over s.
Merle (M), Graying (G), Ticking (T) and Super-Extension (Se) are not recorded as existing in the cat.